Furze is from fyrs the Old English name of this plant.
Other names:
Ajonc (France)
Furze
Gorse
Whin.
Summary:
Gorse (Ulex europaeus)is a perennial, woody, dense spiny evergreen shrub to 4 m high, erect and much-branched with coarse spiny branchlets and leaves reduced to thick spine-tipped phyllodes (although the seedlings have leaves with 3 leaflets). The showy, yellow pea flowers are borne singly and are 15-20 mm long. The hairy seed pod is somewhat compressed and 1-2 cm long.
Native to Europe and originally planted as a hedge, it is now a weed of wasteland and roadsides around Albany. It flowers mainly in autumn and spring.
The seeds remain viable for many years and germination is encouraged by burning.
Description:
Cotyledons:
Two.
First leaves:
Trifoliate.
Leaves:
Mainly modified petioles or phyllodes. True leaves are reduced to scales on mature plants.
In whorled clusters that are fairly uniformly spaced along the branches. Old phyllodes dry off with age but remain attached.
Stipules - None.
Petiole - None.
'Blade' - Usually dark green, narrow and spine-like, 10-40 mm long by 1.5 mm diameter, grooved lengthwise. Tip spined or pointed, edges parallel, base sessile, squarish. Usually hairy.
Stems:
Initially green and turning brown at maturity, erect, much branched, lengthwise ribbed, soft when young and woody and rough when mature, spines to 50 mm long and short branchlets at a wide angle to each other that end in a spine. Low-lying stems may take root where they touch the ground and form layers. Hairy. Forming bushes up to 4000 mm tall and 3000 mm wide.
Flower head:
Sprays of usually single flowers in the leaf axils and very numerous towards the ends of the branches. Flower buds downy.
Flowers:
Bright yellow, pea-type with a coconut like fragrance, 20-25 mm long, on a short stalk that is 5-7 mm long.
Bracts - Shorter than the flower stalk(peduncle), egg shaped, at the base of the calyx..
Ovary -
Perianth -
Sepals - Yellow, divided to the base into 2 lips, 15 mm long. Upper lip has 2 small teeth. Lower lip has 3 teeth. Hairy.
Petals - Longer than calyx. Keel obtuse tipped.
Stamens - United in a complete tube.
Anthers -
Fruit:
Oblong, 2-6 seeded, dark pod, 10-20 mm long by 6 mm wide, oblong and sticking out from the brown calyx bracts. Densely hairy. Explosively ejects seeds when mature.
Seeds:
Green to brown, 3-4 mm long, slightly triangular, smooth, shiny with a yellow to white, fleshy appendages/aril.
Seed size in its native range is similar to that in its exotic range (965). 2102 seeds/m2 shed per year within the canopy (966).
Roots:
Many shallow, woody roots and a few deep roots but no obvious taproot.
Root systems of seedlings are small compared to grasses ().967
Key Characters:
Leaves spiny, simple, entire.
Branchlets spiny.
Calyx scarious, 2 lipped to base.
Staminal tube completely closed around the ovary.
Seeds carunculate.
From J.M. Black.
Biology:
968 have a good review in the Biology of Australian Weeds.
Life cycle:
Perennial. Seed germinates in autumn and spring to mid summer with a peak in spring. The young foliage is soft, and at this stage it is readily grazed by sheep and rabbits. As the plant develops the stems become woody and leaves spine-like. Flowering usually starts in the second season, when they are over 18 months old or it may be delayed until they are 3 years old. Gorse plants can be found in flower at all times of the year. The main flowering period is autumn and through winter with a second main flush in spring. The plants carry relatively little flower in summer. When ripe the seed pod splits open explosively with a sharp crack and seed is thrown a distance of several metres. Mature plants grow for many years (at least 30).
969 has a good model.
970 has a method for determining the age of gorse plants.
In NZ, stems increased in diameter by an average 5 mm/year, in length by 200 mm/year, maximum age was 29 years, maximum height was 7 m and maximum stem diameter 100 mm above ground was 217 mm. Plants mature about 15 years after establishment, with an average stem density of 60,000/ha, average basal area of 51 m2, and litter depth of 55 mm (971).
Physiology:
Mature plants tolerate frost to -200C, young plants are frost sensitive.
Tolerates drought and salt laden winds.
Tolerates burning.
Fixes atmospheric nitrogen. Trifolium species fix 70% more N than gorse (972).
P fertilisation gives increase shoot and root growth and increased nodulation and N and K have little effect (973).
30-40% rainfall reaches the soil under gorse compared to 40-60% under trees (974) and consequently decreased leaching of nutrients (975).
Grows more rapidly when fertiliser N supplied (976).
In low N conditions most of the N in the plant comes from fixed N whereas in high N conditions most of the N comes from root uptake from the soil (977).
Total energy and soluble carbohydrate levels remain constant through the year but nitrogen levels peak in spring which coincides with maximum feeding by phytophagous insects (978).
Surface wax changes after 2-3 months to a more complex structure on juvenile growth of mature plants and on 3 month old seedlings which appears the reduce the uptake of herbicides by mature foliage (979).
Shoots have a high wax content but no obvious wax structures under electron microscopy and a contact angle of droplets of <90 degrees indicating the surface is easy to wet (980).
Photosynthetic to total plant mass ratio decreases as gorse matures and plant weight can be predicted by the maximum bush diameter (981).
In NZ, 19.5 t DM/yr produced with 13.5 t DM/yr green material. In vitro digestibility was 65% DDM in spring and <20% in winter. Levels of N, Ca and Mo in the foliage were adequate for sheep and goat nutrition, but S, K, Cu and Mg were low, and P was very low. Ash concentration averaged 3% of DM (982).
Crude protein peaks at the pre flowering stage at 168 g/kg DM (983).
Generally gorse doesn't affect grass growth but on one alluvial soil it did (984).
Gorse seedlings didn't affect the first year growth of Pinus radiata (985).
Gorse litter increased net mineralization (986).
Soil and microbial C, N and P and macro aggregate stability equal to or greater than native forest (987).
The rhizobia associated with gorse is usually different to those associated with native legumes ().
Reproduction:
By seed and where stems touch the ground and take root.
Honey bees are required to pollinate flowers (988) and other bee species can also pollinate gorse (989).
Flowering times:
Most of the year in SA.
Autumn in WA.
Variable and mainly autumn and July to October in SE Australia.
Seed Biology and Germination:
Germination levels tend to be low and around 10% (990) but sometimes are above 70% (991).
Germination greatest in late summer to early autumn in NZ (992).
Seeds in the centre of the pod have a greater chance of maturation even though all seeds of the same size in the pod. Seed weight did not affect the probability of germination but was positively correlated with seedling weight (993).
Seed from 3 different sources had the same germination responses but differing levels of hard seed (994).
Seed germination was greatest on cultivated plots then areas bared with repeated glyphosate sprays and least on pasture covered areas. However, this didn't lead to significant differences in the amount of gorse that germinated after 6 years at the end of the trials. That is cultivation with a rotary hoe (or spraying) for 6 years reduced the seed bank little more than pasture (992).
Seed collected from blackened pods had a viability of 80% whereas those collected in trays beneath the bushes had 60% viability. There were differences between sites for spring produced seed but not for autumn produced seed (995).
Seed bank numbers
Decline in seed bank approximately exponential when no new seed added and seed buried at 5 cm declines to 10% within 10 years and 1% within 20 years at 2 sites but was much slower at a third site. Losses were nearly all due to germination probably as recovered seed was viable (996). 994 also reports differences in hard seed levels between sites.
25% of seed in the top 10 cm germinated over a 2 year period on areas mechanically cleared whereas seedling numbers declined by 90-95% on an area that burnt and sprayed (997).
Up to 6 million seeds per hectare per year can be produced. (i.e. 600 seeds/m2/yr).
Soil seed bank can exceed 100 million seeds/ha (10,000 seeds/m2).
998 found in Spain in a Mediterranean environment at 2 sites, 645/m2 (95% viable) and 1045/m2 (96.8% viable) seeds were recorded in soil. Most were in the top 5 cm
966 - 2120 seeds/m2 per year are shed.
Hydes & Suckling (1953) - 3050 seeds/m2
Ivens (1978) - 10,000 + 7530/m2
Moss (1959) - 3500/m2
999 - 2600/m2.
Zabkiewicz & Gaskin (1978) - 5446 + 1364/m2
Longevity
No gorse seeds germinated from original surface soil that had been buried under an earth bank for 33 years (Thompson and Band, 1997) Gorse was close by but not verified on the site before burial and only 0.4 m2 was sampled.
Soil bacteria may cause significant seed losses (1000) but 996 in NZ reports that most losses probably due to germination.
Distribution in the soil profile
In Spain, 63% seeds in top 2.5 cm layer of soil, 26% in 2.5-5 cm layer, 10% in 5-10 cm layer and 2% in the 10-15 cm deep soil layer at the first site and a more even distribution of seeds of 30%, 26%, 29%, 15% of seed in the same soil layers at the second site (Puentes and Casal, 1988).
In New Zealand, 71% in the in the top 2.5 cm layer of soil, 14% in the 2.5-5 cm layer and 15% in the 5-10 cm layer of soil (Rolston & Talbot, 1980).
see Gorseseed.xls graph
Burial slightly increases the longevity of seed (996).
Temperature
Pereiras (1984) temps of 80 deg C.
Optimum germination temperature is 15-190C.
Best temperature 15 deg C (991).
Temperature fluctuations may be important in softening impermeable hard seed coats.
994 found little difference between fluctuating and constant temperatures on germination.
Germination rate increased linearly with temperature from 0 deg C to 18 deg C then declined to 26 deg C. At temperatures up to 35 deg C seed imbibed but didn't germinate until cooled. Above 35 deg C viability was lost in wet conditions (994).
Seed can be dry heated to 65 deg C for extraction from the seed pod with minimal loss of viability and 45 deg C with no loss. At 110 deg C viability is lost (Moore 2005 pers com).
Hot water not effective for increasing germination percentage (991).
Light
Germination is not responsive to light () and no red or far red response.
Scarification
Gorse seed germination is about 10%. Mechanical scarification increases germination to 48%. 180-210 minutes in sulfuric acid (36N) gave 81% germination (991).
Potassium nitrate had no effect on germination ().
Burning encourages large germinations of seed after the next rain.
Heat is required to break seed dormancy.
Seeds have a hard coat that protects them. 85% of seed that was 25 years old was still viable.
reports level of hard seed is greatest in the pod seed then the surface seed then the soil seed. This is different to the WA data where soil seed had very low levels of germination (Moore pers com 2005)
Vegetative Propagules:
Stems may take root where they touch the ground.
Hybrids:
Plenus is an ornamental compact variety with double flowers and no seed that is grown in New Zealand.
2 subspecies can be distinguished U. europaeus subspecies europaeuswith small bracteoles and 2n=96 and U. europaeus subspecies latebracteatus with larger bracteoles and 2n=64 chromosomes (1001).
In Europe, there are diploid, tetraploid and hexaploid varieties of Ulex europaeus subsp. europaeus (1002).
1003 has done chromosome counts and 1004 immunological studies and 1005 ploidy levels in the Mediterranean.
Allelopathy:
Population Dynamics and Dispersal:
Spread by seed. Seed is catapulted up to 5000 mm when the pod explosively opens in summer. 40% of seed falls within the canopy of the gorse with a predominance near the edges and little in the centre (966). Birds are a major agent and ants and other animals are minor one in spreading seeds. Earthmoving, road grading and dumping of garden refuse are also important methods of seed dispersal.
Ants spread the seed in Europe (1006).
Seed production in high and up to 6 million seeds may be added to the soil each year. This results in soil seed banks of over 100 million seeds per hectare in old established stands. In NZ, 2120 seeds/m2 were shed per year within the gorse canopy (1007). Only a small proportion (5%) of seed normally germinates each year, which makes control difficult.
Stems may form roots where they contact the soil and increase to size of the infestation.
Competition from ryegrass reduced dry weight of gorse seedlings by 96% (1008).
Displaces Kunzea and Leptospermum as initial post disturbance species (1009).
Successional aspects in heathlands have been studied. (190, 1010, 1011).
If dead material excluded and other species included then gorse self thins at the theoretical slope of -3/2 (1012).
Gorse is a primary coloniser and disappeared after 20-30 years in the UK (1013).
When fire and grazing animals are removed, native forest usually regenerates with time in NZ (1014).
Origin and History:
Western Europe and North Africa.
Probably introduced soon after settlement and records of it go back to 1803.
Introduced to many areas as a hedge.
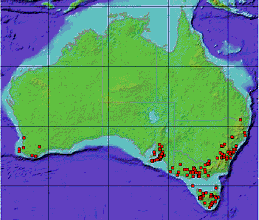
Distribution:
NSW, QLD, SA, TAS, VIC, WA.
Occurs in all farmed areas of Tasmania and is most prevalent in the Midlands.
Major infestations are in Tasmania, southern and central Victoria and New Zealand (1015). 1.84% of NZ south island agricultural land in covered by gorse (1016).
It is also in British Columbia (1017), Canada noxious weed (1018), Chile (1018, 1019), 15000 ha in Hawaii (1020, 1021, 1022), Ireland (1023), Italy (1024), Poland (1025), Reunion Island (1026), Scotland (1027), USA (1018)
1015 has a survey of 37 countries.
Courtesy Australia's Virtual Herbarium
Habitats:
Prefers full sun but tolerates deep shade with a survival of 74% (1028).
Tolerates a wide range of habitats (1029).
Climate:
Temperate. Mediterranean.
Most abundant in frost free areas in higher rainfall areas with 600-900 mm annual rainfall.
1030 has world distribution based on climate.
Gorse consumes more water than pasture but 63 mm/year less than a pine plantation including the establishment phase (1031).
Soil:
Grows well on most soils from light sands to heavy clays.
Most abundant on fertile acidic soils and tends to avoid the calcareous soils.
Can grow on very nitrogen deficient soils because it fixes atmospheric N.
Rarely dominates on soils with a maximum P absorption level of <70 ugP/g soil and prefers soils within the 70-700 range. At lower or higher levels other species dominate (1032).
Plant Associations:
Most common on creek lines, roadsides, ungrazed areas and forest margins.
Can coexist with Scotch broom (Cytisus scoparius) but tends to prefer drier sites (1018).
Significance:
In NZ, a 50% control would result in a direct benefit of more than $18 million per year and would cost beekeepers $1.6 million per year (1033). The benefit:cost ratio of biocontrol for gorse in NZ is 12:1 if successful (1034).
1035 has the economics of control in plantations.
Beneficial:
Utilised by sheep for feed when young or when other food is not available.
If managed properly it can be used for hedges, wind breaks and erosion control (1036, 1037).
It is a plentiful source of pollen for bees in autumn and especially in early spring when it is most needed (1037). Bumble bees collect pollen in England (1038, 1039, 1040).
The seed is the source of chemicals used for blood grouping and agglutination procedures and a gorse lectin is used for immunohistochemical tests (1041).
Ulex europaeus agglutinin I (UEA1) has been used as a ligand for targeted oral vaccine delivery to M cells (antigen-presenting cells) (1042).
Extracts have action against Herpes simplex virus (1043, 1044) and antibacterial properties (1045) and antifungal properties (1046).
Some endophytes of gorse have antibiotic properties (1047).
Provides habitat for endangered warbler in England (1048).
Considered for pyrolysis conversion as a source of energy in Britain (1049).
Calorific value as fuel is highest in summer and lowest in autumn and spring (1050). Values of 7500-7600 kJ/kg recorded by 1051 and 19000 kJ/kg by 1052 with yields of 20 t/ha (1053).
It is associated with Aries in astrology and typifies cheerfulness in adversity.
Used for dye.
Fixes nitrogen (1054).
Detrimental:
Weed of pasture, roadsides, plantations (1055), forest margins, mountain forests (1056), urban bushland (1057), mine dumps and disturbed areas.
It is an invasive and competitive plant in grazing land and pasture.
It often takes over forest areas after they have been logged and prevents the re establishment of native species. Seedlings reduce the growth of Pinus radiata (1058) but 1059 and 1060 reports little effect of gorse on Pinus radiata growth at one site and reduced stem volumes at a drier site (1061). Threatens native vegetation in NZ (1062) and ecosystems in Chile (1063) and endangered ecosystems in British Columbia (1017).
Forms impenetrable thickets impeding the movement of people and stock and excluding most other plant species (1018).
Occasionally it will replace native plants in parks and reserves.
Spines may injure stock and humans.
Very severe fire hazard. Two fires in NZ spread at 4400 m/hr with an intensity of 60000 kw/m2 and 3400 m/hr and 25000 kw/m2 which is pretty hot (1064). Poses a fire hazard in pine plantations which can be alleviated by sowing pastures and grazing (1065).
Harbours rabbits and vermin.
Acidifies the soil.
Listed as a 'Weed of National Significance' and a "Garden thug'.
Dwarf Furze (Ulex minor) is smaller and has naturalised in NZ but has not been found in Australia yet.
Plants of similar appearance:
Broom (Cytisus scoparius) is not prickly.
Montpellier Broom (Genista monspessulana) is not prickly and has trifoliate leaves.
Miscellaneous
1.5-5.4% of gross margin spent on weed control in NZ.
Fact-sheet hand-outs of regionally-based farm-sized trials and short, concise articles in farming magazines were the favoured means of technology transfer (1066).
Trampling by people especially in winter caused an increase in goorse dominance (1067).
References:
Auld, B.A. and Medd R.W. (1992). Weeds. An illustrated botanical guide to the weeds of Australia. (Inkata Press, Melbourne). P170. Photo.
Black, J.M. (1965). Flora of South Australia. (Government Printer, Adelaide, South Australia). P449-450. Diagram.
Blood, K. (2001). Environmental weeds: a field guide for SE Australia. (CH Jerram & Associates, Australia). P82-83. Photos.
Hussey, B.M.J., Keighery, G.J., Cousens, R.D., Dodd, J. and Lloyd, S.G. (1997). Western Weeds. A guide to the weeds of Western Australia. (Plant Protection Society of Western Australia, Perth, Western Australia). P162-163. Photo.
Hyde-Wyatt, B.H. and Morris, D.I. (1980) The Noxious and Secondary Weeds of Tasmania. (Tasmanian Department of Agriculture, Hobart, Tasmania). P50-51. Diagrams.
Lamp, C. and Collet, F. (1990). A Field Guide to Weeds in Australia. (Inkata Press, Melbourne).
Lazarides, M. and Hince, B. (1993). CSIRO handbook of economic plants of Australia. (CSIRO, Melbourne). #1245.1.
Moore, J.H. and Wheeler, J.R. (2002). Southern Weeds and their Control. Photos.
Muyt, A. (2001). Bush Invaders of South-East Australia: a guide to the identification and control of environmental weeds found in South East Australia. (R.G and F.J. Richardson, Australia). P224-226. Photos.
Parsons, W.T. and Cuthbertson, E.G. (1992). Noxious weeds of Australia. (Inkata Press, Melbourne). P480-484. Photos.
Acknowledgments:
Collated by HerbiGuide. Phone 08 98444064 or www.herbiguide.com.au for more information.